








Views: 24
“Como lo hemos estado viendo en ediciones anteriores, el agua es un componente esencial en la vida de cualquier organismo y sobretodo en las plantas. No solo por ser el medio de transporte de elementos nutricionales, sino por ser un termorregulador de las células que proporcionan turgencia y vigor.”
Dr. Luis Alberto Lightbourn Rojas PhD.
Presidente Instituto Lightbourn Research. Systems Biology.
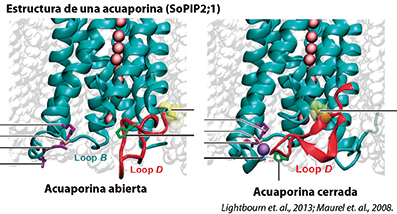
El agua fluye a través de las membranas de las células vegetales y se regula básicamente por dos mecanismos que implican la cantidad de acuaporinas y la actividad de las mismas proteínas. Ante la presencia de diversos agentes que promueven la oxidación de las membranas como los nitratos y sulfatos, provocan una reducción en el transporte del agua debido a que afectan de manera directa la actividad acuaporínica.
El bloqueo de estas, puede reducirse entre un 30 a un 40% de la conductividad hidráulica de la raíz. Para evitarlo, se requiere el uso de agentes que provoquen un efecto reversible a los daños causados por el uso de nitratos y sulfatos, como el níquel, que en balance con el selenio, titanio y zinc, despolariza la membrana sosteniendo el equilibrio iónico Gibbs- Donnan, así como el transporte vacuolar, en especial en condiciones extremas de temperatura ya que se evita la coagulación intravascular diseminada en el cambium, con el consecuente bloqueo acropetálico de los nutrientes.
La germinación de las semillas depende de la cantidad de agua que esta pueda adquirir, lo cual se correlaciona con la expresión de acuaporinas en esta etapa de desarrollo, en particular con las PIPs. También se encuentra específicamente en células vasculares, de las que se sugiere, participan en el transporte de savia entre los diversos órganos de la planta.
La expresión del TIP1, 1 se ha asociado con el alargamiento de las raíces, hipocólitos y hojas. Acumulándose en los transcritos mediante la hormona de crecimiento ácido giberélico que está asociada a la Ribofuranosilzeatina en relación al balance de níquel, selenio y titanio.
El nitrógeno es un elemento esencial para el desarrollo de las plantas y uno de los más utilizados en la nutrición vegetal. Las acuaporinas PIPS, TIPs y NIPs, participan en la transportación de compuestos nitrogenados como el amoniaco, así como el ácido anhídrido carbónico, tanto en la membrana citoplasmática como en la membrana de los cloroplastos. Por lo que influyen directamente en los procesos de fotosíntesis.
El calcio y el boro son indispensables para el desarrollo de la pared celular, por lo que las deficiencias del último mencionado, muestran una reducción significativa del crecimiento de tallos y raíces, como en la formación de flores.
Estas deficiencias muestran una sobreexpresión de acuaporinas NIPs que transportan este elemento a través de las raíces para el desarrollo óptimo de la planta.
La expresión de PIP1 ha demostrado poder incrementar la obtención de boro hasta en un 30%. En el caso del silicio, ayuda a mejorar el sistema inmunológico natural de las plantas ante el ataque de diversos patógenos.
Algunas acuaporinas TIPs tienen la capacidad de transportar glicerol. Y recientes investigaciones reportan que una inhibición en estas acuaporinas presenta una reducción significativa en el crecimiento sano de las raíces.
Efectos del estrés en las acuaporinas
Las acuaporinas pueden afectarse por distintos tipos de estrés como lo son las deficiencias de algunos nutrientes y micronutrientes esenciales para el desarrollo de las plantas, por ejemplo: deficiencia de nitrógeno, fosforo o azufre que reducen el transporte de agua en las raíces. Sin embargo esta reducción puede ser reversible aunque puede tardar poco más de 24 horas, por lo que la planta podría mantener daños irreversibles durante su proceso de deshidratación.
En lo que corresponde al calcio, reduce significativamente la transcripción de acuaporinas, perdiendo la densidad de las mismas membranas promoviendo un menor curso de componentes que mueven a estas proteínas.
Sin dejar de mencionar que es común la deficiencia del calcio cuando el ratio Na/Ca es elevado en el agua del suelo. Así los iones de sodio compiten con los iones del calcio por los sitios de enlace con las membranas disminuyendo su obtención.
El estrés por frio, provoca que se reduzca el fuljo de savia y la conductividad hidráulica en las raíces en poco tiempo en diversos tipos de cultivo perennes, lo cual conlleva a problemas de hidratación y transporte de solutos en toda la planta.
Lo que merma la actividad de las acuaporinas, que es mediado por radicales libres producidos durante este tipo de estrés y puede llegar a dañar el ADN o a las acuaporinas modificando su estructura o actividad.
La anoxia (ausencia de oxigeno) es provocada por las inundaciones, riego excesivo o compactación de los suelos inducida por las diversas prácticas agrícolas inadecuadas, que es una de las causas principales.
La anoxia reduce el pH de las células radiculares que con la aplicación de fertilizantes ácidos se vuelve más severo. Lo que hace que disminuya la conductividad hidráulica asociada al cierre de las acuaporinas PIP causada por el exceso de protones en el citoplasma.
El exceso de salinidad es una de las causas más preocupantes de estrés en los suelos de cultivo a nivel mundial, ya que es uno de los factores que limita a las plantas para la obtención de agua y nutrientes de forma correcta, que aminora el crecimiento y producción total de las cosechas.
La salinidad del suelo y el exceso de riegos en el logro de la solubilidad de fertilizantes salinos acortan esa conductividad en las células vegetales por diversas causas entre las que se encuentran: la regulación genética y su impacto directo en la actividad de las acuaporinas.
Se ha demostrado que dicha salinidad aminora la transcripción de las principales acuaporinas TIPs y PIPs en un 60% A 70% en poco tiempo. Así como se ha observado la redistribución de las acuaporinas durante los efectos del estrés salino, por ejemplo en las TIPs se colocan invaginaciones producidas por vacuolas; mientras que en las PIPs, se encuentran en estructuras intracelulares que inhiben su traslado hacia la membrana citoplasmática e incluso algunas acuaporinas se degradan en proteosomas.
Se dice que los problemas por salinidad se pueden disminuir con calcio, aunque al aplicar cloruro de calcio, puede percibirse que la cantidad de acuaporinas PIPs se minimiza y la cantidad de estas se ve restablecido con un porcentaje considerable.
Aunque cuando a las células vegetales se les aplica KCL y CaCl2 se muestra una disminución de hasta 4 veces de la conductividad comparado con células control debido a los efectos adversos.
En contraste, se nota un incremento dos veces más en esta conductividad en la célula cuando los iones de cloro son sustituidos. Por lo tanto, los iones Cl provocan efectos deletéreos en la producción y actividad de las acuaporinas.
De este modo, se ha evitado con la aplicación de calcio con base coloidal, que actúa de forma natural y compatible mediante una cascada de quinasas para producir y activar eficientemente a las acuaporinas, a su vez promoviendo la activación de calmodulinas, que en conjunto con tetra péptidos activos de cisteína-glicina-prolina-cisteína, que continúan una cascada de señales a través de ferredoxina y epimerasas, que en sinergia con los elementos N, P, K, Ca, B, Zn, Mg y Fe logran la producción y estabilización de tubulinas.
Al aplicar nutrientes compatibles con la naturaleza coloidal de suelos y estructuras vegetales, el productor agrícola garantiza de esta forma restablecer el transporte de agua y obtención de nutrientes de la planta a nivel celular, en la que las acuaporinas juegan un papel esencial para favorecer la estructura y producción de nuevas células vegetales, detonando así, el crecimiento abundante de los cultivos.
Bibliografía:
Barrieu F, Morrillon R, Chrispeels M J, 2000. Modulation of aquaporin gene expression in Arabidopsis leads to altered membrane water permeability. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Bhardwaj R, Sharma I, Kanwar M, Sharma R, Handa N, Kaur H, Kapoor D, Poonam, 2013. Aquaporins: role under salt stress in plants. In: Ecophysiology and response of plants under salt stress. Ahmad P, Azooz M M, Prasad M N V, Springer
Carvajal M, Martínez-Ballesta M C, Martínez V, 2000. The response of plants to salinity involves root and water channels. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Chaumont F, Tyerman S D, 2014. Aquaporins: highly regulated channels controlling plant water relations. Plant Physiology 164:1600-1618
Chaumont F, Van Wilder V, Felter C, Barrieu F, Chrispeels M J, 2000. Characterization of plasma membrane MIP proteins in maize. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Chevalier A S, Chaumomnt F, 2015. Trafficking of plant plasma membrane aquaporins: multiple regulation levels and complex sorting signals. Plant Cell Physiology 56(5):819-829
Cochard H, Venisse J-S, Barigah T S, Brunel N, Herbette S, Guilliot A, Tyree M T, Sakr S, 2007. Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiology 143:122-133
Cooper G J, Dixit R, Nasrallah J, Boron W F, 2000. The permeability of MIPs to gases. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Gerbeau P, Javot H, Lauvergeat V, Martin-Laurent F, Güςlü J, Santoni V, Tournaire C, Ripoche P, Maurel C, 2000. Physiological and genetic analysis of plasma membrane aquaporin functions in Arabidopsis. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Luu D-T, Maurel C, 2013. Aquaporin trafficking in plant cells: an emerging membrane-protein model. Traffic 14:629-635
“Como lo hemos estado viendo en ediciones anteriores, el agua es un componente esencial en la vida de cualquier organismo y sobretodo en las plantas. No solo por ser el medio de transporte de elementos nutricionales, sino por ser un termorregulador de las células que proporcionan turgencia y vigor.”
Dr. Luis Alberto Lightbourn Rojas PhD.
Presidente Instituto Lightbourn Research. Systems Biology.
El agua fluye a través de las membranas de las células vegetales y se regula básicamente por dos mecanismos que implican la cantidad de acuaporinas y la actividad de las mismas proteínas. Ante la presencia de diversos agentes que promueven la oxidación de las membranas como los nitratos y sulfatos, provocan una reducción en el transporte del agua debido a que afectan de manera directa la actividad acuaporínica.
El bloqueo de estas, puede reducirse entre un 30 a un 40% de la conductividad hidráulica de la raíz. Para evitarlo, se requiere el uso de agentes que provoquen un efecto reversible a los daños causados por el uso de nitratos y sulfatos, como el níquel, que en balance con el selenio, titanio y zinc, despolariza la membrana sosteniendo el equilibrio iónico Gibbs- Donnan, así como el transporte vacuolar, en especial en condiciones extremas de temperatura ya que se evita la coagulación intravascular diseminada en el cambium, con el consecuente bloqueo acropetálico de los nutrientes.
La germinación de las semillas depende de la cantidad de agua que esta pueda adquirir, lo cual se correlaciona con la expresión de acuaporinas en esta etapa de desarrollo, en particular con las PIPs. También se encuentra específicamente en células vasculares, de las que se sugiere, participan en el transporte de savia entre los diversos órganos de la planta.
La expresión del TIP1, 1 se ha asociado con el alargamiento de las raíces, hipocólitos y hojas. Acumulándose en los transcritos mediante la hormona de crecimiento ácido giberélico que está asociada a la Ribofuranosilzeatina en relación al balance de níquel, selenio y titanio.
El nitrógeno es un elemento esencial para el desarrollo de las plantas y uno de los más utilizados en la nutrición vegetal. Las acuaporinas PIPS, TIPs y NIPs, participan en la transportación de compuestos nitrogenados como el amoniaco, así como el ácido anhídrido carbónico, tanto en la membrana citoplasmática como en la membrana de los cloroplastos. Por lo que influyen directamente en los procesos de fotosíntesis.
El calcio y el boro son indispensables para el desarrollo de la pared celular, por lo que las deficiencias del último mencionado, muestran una reducción significativa del crecimiento de tallos y raíces, como en la formación de flores.
Estas deficiencias muestran una sobreexpresión de acuaporinas NIPs que transportan este elemento a través de las raíces para el desarrollo óptimo de la planta.
La expresión de PIP1 ha demostrado poder incrementar la obtención de boro hasta en un 30%. En el caso del silicio, ayuda a mejorar el sistema inmunológico natural de las plantas ante el ataque de diversos patógenos.
Algunas acuaporinas TIPs tienen la capacidad de transportar glicerol. Y recientes investigaciones reportan que una inhibición en estas acuaporinas presenta una reducción significativa en el crecimiento sano de las raíces.
Efectos del estrés en las acuaporinas
Las acuaporinas pueden afectarse por distintos tipos de estrés como lo son las deficiencias de algunos nutrientes y micronutrientes esenciales para el desarrollo de las plantas, por ejemplo: deficiencia de nitrógeno, fosforo o azufre que reducen el transporte de agua en las raíces. Sin embargo esta reducción puede ser reversible aunque puede tardar poco más de 24 horas, por lo que la planta podría mantener daños irreversibles durante su proceso de deshidratación.
En lo que corresponde al calcio, reduce significativamente la transcripción de acuaporinas, perdiendo la densidad de las mismas membranas promoviendo un menor curso de componentes que mueven a estas proteínas.
Sin dejar de mencionar que es común la deficiencia del calcio cuando el ratio Na/Ca es elevado en el agua del suelo. Así los iones de sodio compiten con los iones del calcio por los sitios de enlace con las membranas disminuyendo su obtención.
El estrés por frio, provoca que se reduzca el fuljo de savia y la conductividad hidráulica en las raíces en poco tiempo en diversos tipos de cultivo perennes, lo cual conlleva a problemas de hidratación y transporte de solutos en toda la planta.
Lo que merma la actividad de las acuaporinas, que es mediado por radicales libres producidos durante este tipo de estrés y puede llegar a dañar el ADN o a las acuaporinas modificando su estructura o actividad.
La anoxia (ausencia de oxigeno) es provocada por las inundaciones, riego excesivo o compactación de los suelos inducida por las diversas prácticas agrícolas inadecuadas, que es una de las causas principales.
La anoxia reduce el pH de las células radiculares que con la aplicación de fertilizantes ácidos se vuelve más severo. Lo que hace que disminuya la conductividad hidráulica asociada al cierre de las acuaporinas PIP causada por el exceso de protones en el citoplasma.
El exceso de salinidad es una de las causas más preocupantes de estrés en los suelos de cultivo a nivel mundial, ya que es uno de los factores que limita a las plantas para la obtención de agua y nutrientes de forma correcta, que aminora el crecimiento y producción total de las cosechas.
La salinidad del suelo y el exceso de riegos en el logro de la solubilidad de fertilizantes salinos acortan esa conductividad en las células vegetales por diversas causas entre las que se encuentran: la regulación genética y su impacto directo en la actividad de las acuaporinas.
Se ha demostrado que dicha salinidad aminora la transcripción de las principales acuaporinas TIPs y PIPs en un 60% A 70% en poco tiempo. Así como se ha observado la redistribución de las acuaporinas durante los efectos del estrés salino, por ejemplo en las TIPs se colocan invaginaciones producidas por vacuolas; mientras que en las PIPs, se encuentran en estructuras intracelulares que inhiben su traslado hacia la membrana citoplasmática e incluso algunas acuaporinas se degradan en proteosomas.
Se dice que los problemas por salinidad se pueden disminuir con calcio, aunque al aplicar cloruro de calcio, puede percibirse que la cantidad de acuaporinas PIPs se minimiza y la cantidad de estas se ve restablecido con un porcentaje considerable.
Aunque cuando a las células vegetales se les aplica KCL y CaCl2 se muestra una disminución de hasta 4 veces de la conductividad comparado con células control debido a los efectos adversos.
En contraste, se nota un incremento dos veces más en esta conductividad en la célula cuando los iones de cloro son sustituidos. Por lo tanto, los iones Cl provocan efectos deletéreos en la producción y actividad de las acuaporinas.
De este modo, se ha evitado con la aplicación de calcio con base coloidal, que actúa de forma natural y compatible mediante una cascada de quinasas para producir y activar eficientemente a las acuaporinas, a su vez promoviendo la activación de calmodulinas, que en conjunto con tetra péptidos activos de cisteína-glicina-prolina-cisteína, que continúan una cascada de señales a través de ferredoxina y epimerasas, que en sinergia con los elementos N, P, K, Ca, B, Zn, Mg y Fe logran la producción y estabilización de tubulinas.
Al aplicar nutrientes compatibles con la naturaleza coloidal de suelos y estructuras vegetales, el productor agrícola garantiza de esta forma restablecer el transporte de agua y obtención de nutrientes de la planta a nivel celular, en la que las acuaporinas juegan un papel esencial para favorecer la estructura y producción de nuevas células vegetales, detonando así, el crecimiento abundante de los cultivos.
Bibliografía:
Barrieu F, Morrillon R, Chrispeels M J, 2000. Modulation of aquaporin gene expression in Arabidopsis leads to altered membrane water permeability. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Bhardwaj R, Sharma I, Kanwar M, Sharma R, Handa N, Kaur H, Kapoor D, Poonam, 2013. Aquaporins: role under salt stress in plants. In: Ecophysiology and response of plants under salt stress. Ahmad P, Azooz M M, Prasad M N V, Springer
Carvajal M, Martínez-Ballesta M C, Martínez V, 2000. The response of plants to salinity involves root and water channels. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Chaumont F, Tyerman S D, 2014. Aquaporins: highly regulated channels controlling plant water relations. Plant Physiology 164:1600-1618
Chaumont F, Van Wilder V, Felter C, Barrieu F, Chrispeels M J, 2000. Characterization of plasma membrane MIP proteins in maize. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Chevalier A S, Chaumomnt F, 2015. Trafficking of plant plasma membrane aquaporins: multiple regulation levels and complex sorting signals. Plant Cell Physiology 56(5):819-829
Cochard H, Venisse J-S, Barigah T S, Brunel N, Herbette S, Guilliot A, Tyree M T, Sakr S, 2007. Putative role of aquaporins in variable hydraulic conductance of leaves in response to light. Plant Physiology 143:122-133
Cooper G J, Dixit R, Nasrallah J, Boron W F, 2000. The permeability of MIPs to gases. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Gerbeau P, Javot H, Lauvergeat V, Martin-Laurent F, Güςlü J, Santoni V, Tournaire C, Ripoche P, Maurel C, 2000. Physiological and genetic analysis of plasma membrane aquaporin functions in Arabidopsis. In: Molecular biology and physiology of water and solute transport. Hohmann S, Nielsen S.
Luu D-T, Maurel C, 2013. Aquaporin trafficking in plant cells: an emerging membrane-protein model. Traffic 14:629-635
Martínez-Ballesta M C, Cabañero F, Olmos E, Periago P M, Maurel C, Carvajal M, 2008. Two different effect of calcium on aquaporins in salinity-stressed pepper plants. Planta 228:15-25
Martínez-Ballesta M C, Silva C, López-Berenguer C, Cabañero F J, Carvajal M, 2006. Plant aquaporin: new perspectives on water and nutrient uptake in saline environment
Maurel C, Verdoucq L, Luu D-T, Santoni V, 2008. Plant aquaporins: membrane channels with multiple integrated functions. The Annual Review of Plant Biology 59:595-624
Quian Z-J, Song J-J, Chaumont F, Ye Q, 2015. Differential response of plasma membrane aquaporins in mediating water transport of cucumber seedlings under osmotic and salt stress. Plant, Cell and Environment
Articulos relacionados:
 Estructura y Función de las Acuaporinas en las Células Vegetales
Estructura y Función de las Acuaporinas en las Células Vegetales
 Estructura y Mecanismos de regulación molecular de las acuaporinas
Estructura y Mecanismos de regulación molecular de las acuaporinas
 La biotecnología en algodón
La biotecnología en algodón
 ¡Haciendo preguntas efectivas!. Tercera parte
¡Haciendo preguntas efectivas!. Tercera parte
 BlueDrip
BlueDrip
 Innovaciones para vida útil de frutas y hortalizas
Innovaciones para vida útil de frutas y hortalizas
 Adopción de biotecnología de cultivos Parte 1
Adopción de biotecnología de cultivos Parte 1
 Semillas y biotecnología: verdades y necesidades
Semillas y biotecnología: verdades y necesidades


